 实验热线:4006991663
实验热线:4006991663- RNA-蛋白/RNA互作
- RNA pull down

- circRNA pull down
- miRNA pull down
- RIP
- DNA-蛋白互作
- DNA pull down
- ChIP (染色质免疫共沉淀)
- 化合物-蛋白互作
- 化合物pull down
- 表达细胞株
- 基因表达稳转株
- 抗体表达稳转株(CHO细胞)
- 敲除细胞株
- 基因敲除单克隆株(Crispr-cas9)
实验热线:4006991663如果没有抗原的存在,人类免疫系统可以产生10多种12不同的抗体和多达10种18对外来抗原的反应。哺乳动物免疫系统已经进化到通过基因片段连接和体细胞超突变等机制产生比基因更多的抗体(图1)。这些机制驱动抗体多样性,使免疫系统能够快速检测和中和病原体,并微调免疫反应。
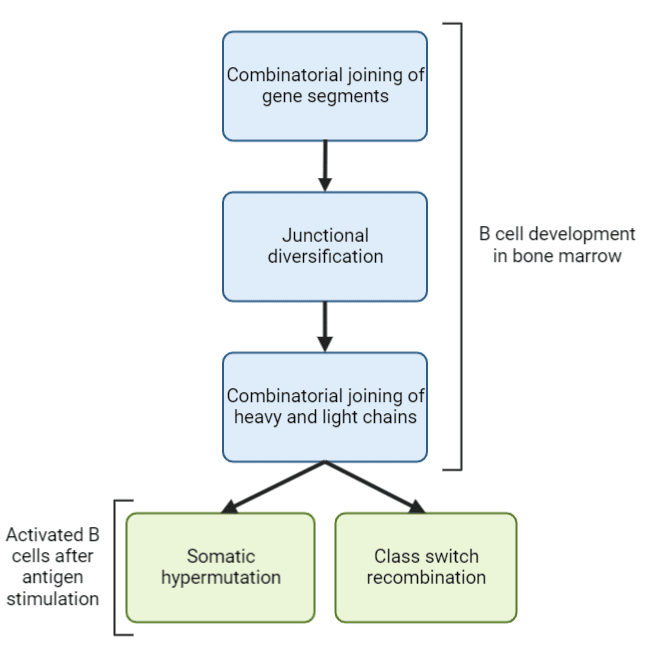
图一:产生抗体多样性的主要机制。
抗体基因座包含各种基因片段,这些基因片段在发育中的B细胞中重新排列以产生高度可变的抗体库。轻链可变区(LC)由两个基因片段组装而成——一个长可变(V)片段和一个短连接(J)片段(图2)。重链(HC)可变区由V区段、J区段和多样性(D)区段组装而成。
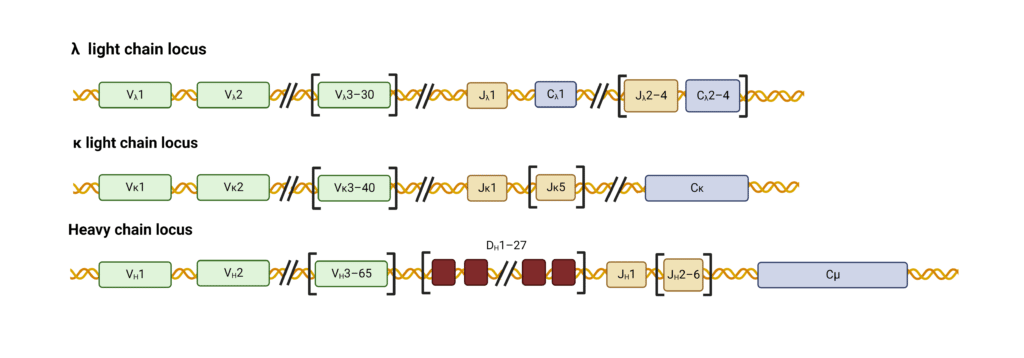
图二:人类重链和轻链基因座中基因片段的组织。
V、D和J区段的重组和连接产生了功能可变的免疫球蛋白区域(图3)。对于人κ LCs,40 V片段可以与5 J片段中的任何一个结合,导致由该库编码的200种可能的组合。人类λ LCs的基因包含30 V区段和4 J区段,导致120个可能的可变λ区域。HC含有65 V片段,可与6 J和27 D片段中的任何一个结合,形成大约11,000个可能的可变HC区。
称为位点特异性重组的过程介导V(D)J复合。染色体DNA双链断裂是由RAG重组酶引入的。这种酶复合物在每个V、D和J片段侧翼的保守序列处结合并切割DNA。选定的V、D和J片段重新排列并连接形成V(D)J外显子。DNA末端被DNA修复酶修复,导致基因片段的缺失或倒位。
在重组过程中,核苷酸经常在连接位点丢失或插入,导致移码突变。这个过程被称为交界多样化在可变区引入了额外水平的抗体多样性,特别是在第三互补决定区(CDR3)。形成的V(D)J外显子被转录并翻译成功能性HC或LC(图3)。
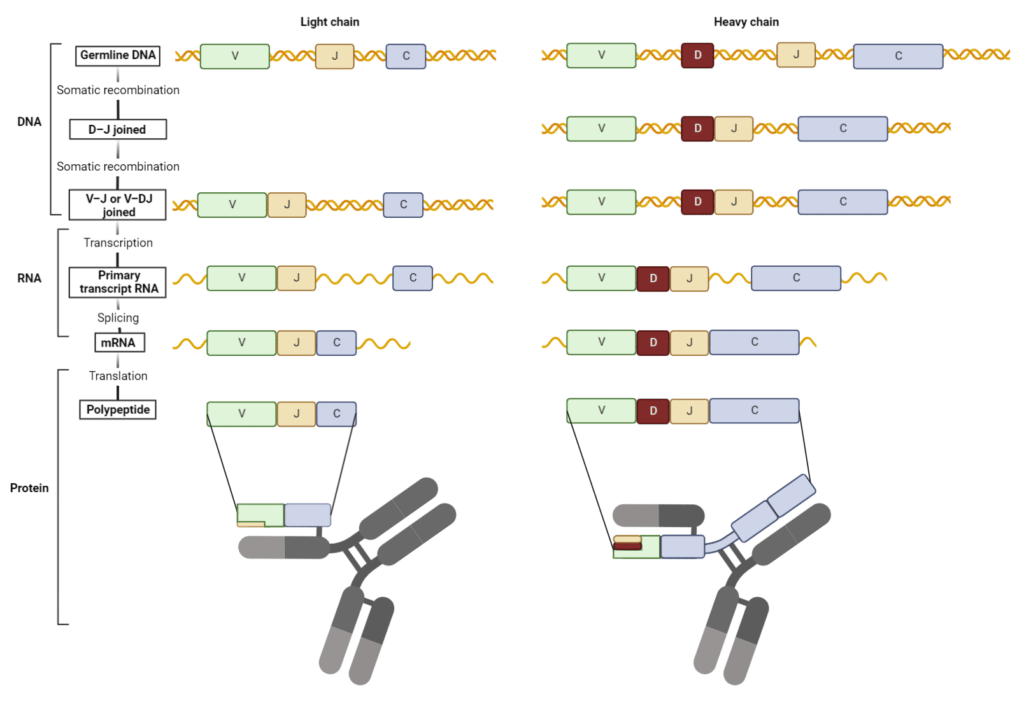
图3:V(D)J重组形成重链和轻链的可变区
在单个B细胞中,任何可能的HC都可以重组,并与任何可能的LC一起产生。由于两个H+L链都在抗原结合位点提供抗原特异性,这导致超过300万个独特的抗体。组合多样性通过区段连接和H+LC配对的不同组合来实现抗原结合位点的结合。
躯体超突变(SHM)在V(D)J重组后提供了额外水平的抗体多样性。由体细胞超突变引起的点突变增加了可变区的变异,发生频率是其他基因突变的一百万倍。双链DNA断裂被引入可变区,引发DNA损伤反应途径,促进易错修复并产生突变抗体。
SHM有助于抗体对被称为亲和力成熟(图4)。一些免疫球蛋白突变体比其他突变体更好地结合抗原,而一些突变体产生非生产性重排。的突变框架区域可变结构域的抗体倾向于被选择对抗,因为它们不增强抗原结合和改变基本的抗体结构。增强抗原结合的突变倾向于聚集在CDR区域。高突变的B细胞经历了一个选择性过程生发中心其中B细胞以亲和力依赖的方式竞争各种信号,从而胜过亲和力较低的B细胞克隆。选择强亲和力的结合物并增殖和成熟为抗体分泌细胞,而较低亲和力的克隆通过细胞凋亡。
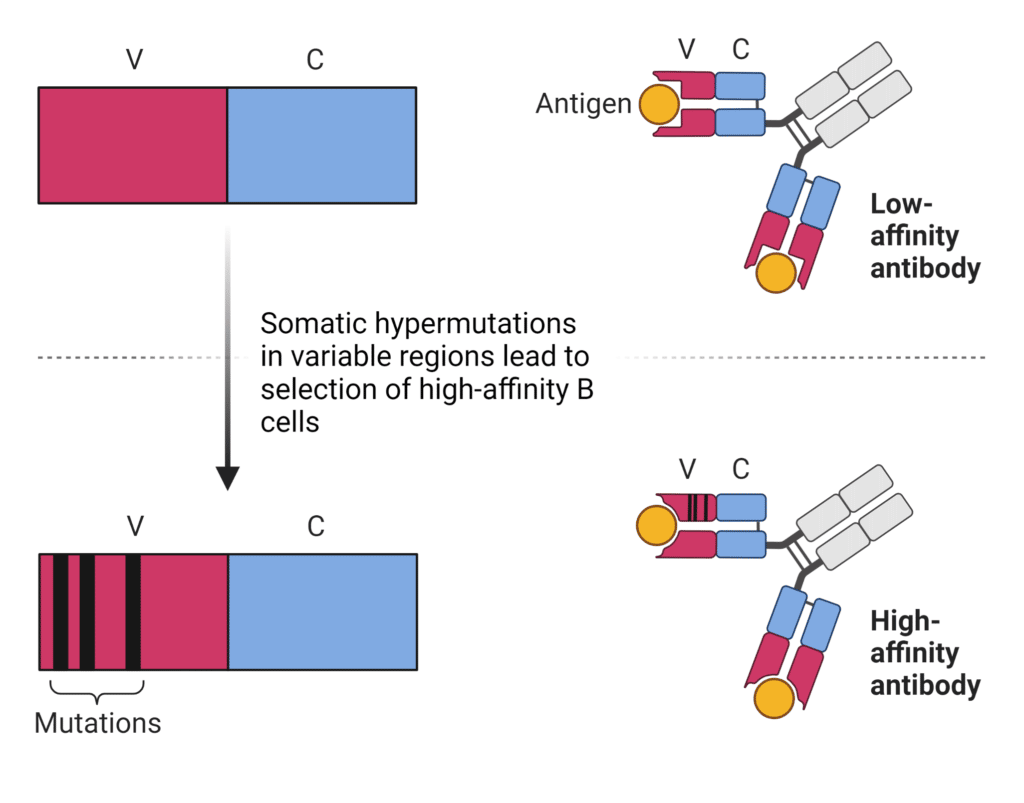
图4:体细胞超突变导致更高亲和力的抗体。
在B细胞发育过程中,细胞在生成一个抗体类别对另一个人来说,这个过程叫做类别转换重组(CSR)。IgM是B细胞合成的第一种免疫球蛋白,B细胞作为B细胞受体(BCR)插入质膜。当B细胞离开骨髓时,它们开始合成嵌在质膜内的IgM和IgD分子。一旦这些B细胞通过BCR与抗原相互作用并结合,就会导致膜结合IgM转变为可溶性IgM。随着免疫反应的进行,发生几个类别转换事件以产生IgG、IgE或IgA抗体,通过记忆B细胞产生二次抗体反应。
HC的恒定区决定了抗体的种类。因此,企业社会责任通过以下方式发生染色体内缺失HC恒定区内的开关区。CSR不同于V(D)J重组,因为CSR发生在抗原刺激后,由辅助T细胞激活,并涉及识别DNA上不同侧翼序列的不同酶。CSR仅改变恒定区而不影响抗原结合位点。因此,CSR将抗原结合位点分布在各种抗体类别中,并提供效应子反应和生物功能的多样性。
辉骏生物基于高精度、高分辨率质谱设备,结合自主研发的测序技术平台,能够实现对抗体氨基酸序列的精准解读。同时,我们有自建的高通量表达验证平台,能够对已测得的抗体序列进行高通量表达验证,从而保证交付抗体的活性。
专业提供抗体测序技术服务,抗体从头测序服务,单克隆抗体从头测序服务,蛋白质测序,抗体表达等多类抗体实验外包。辉骏专注于品质和创新,以极具竞争力的价格和最全面的服务,满足您的实验需求,目前已经与国内外多家知名企业建立了合作关系。


上一篇:什么是质谱(MS)蛋白质测序?
©2011-2026 广州辉骏生物科技股份有限公司 主营业务:RNA pull down DNA pull down GST pull down CoIP LC-MS/MS TAP-MS 抗体测序 版权所有 粤ICP备19156356号 | 网站地图


